Строение и болезни конечностей лошадей. Макроэволюция
Одним из самых известных и наиболее изученных из является филогенетический ряд современных однопалых копытных. Множественные палеонтологические находки и выявленные переходные формы создают научную доказательную базу данного ряда. Описанный русским биологом Владимиром Онуфриевичем Ковалевским еще в 1873 году филогенетический ряд лошади и сегодня остается «иконой» эволюционной палеонтологии.
Эволюция через века
В эволюции филогенетические ряды - это последовательно сменявшие друг друга переходные формы, приведшие к формированию современных видов. По количеству звеньев ряд может быть полным или частичным, однако наличие последовательных переходных форм является обязательным условием их описания.
Филогенетический ряд лошади относят к доказательствам эволюции именно благодаря наличию таких последовательных форм, сменяющих друг друга. Множественность палеонтологических находок наделяет его высокой степенью достоверности.
Примеры филогенетических рядов
Ряд лошадей не единственный среди описанных примеров. Хорошо изучен и имеет высокую степень достоверности филогенетический ряд китов и птиц. А спорным в научных кругах и наиболее используемым при различных популистских инсинуациях является филогенетический ряд современных шимпанзе и человека. Споры по поводу недостающих здесь промежуточных звеньев не утихают в научной среде. Но сколько бы ни было точек зрения, бесспорным остается значение филогенетических рядов как доказательств эволюционной приспособляемости организмов к изменяющимся условиям окружающей среды.

Связь эволюции лошадей с окружающей средой
Множественные исследования палеонтологов подтвердили теорию О. В. Ковалевского о тесной взаимосвязи изменений в скелете предков лошадей с изменениями окружающей среды. Меняющийся климат приводил к уменьшению лесных массивов, и предки современных однопалых копытных приспосабливались к условиям жизни в степях. Необходимость в быстром передвижении провоцировала модификации строения и количества пальцев на конечностях, изменение скелета и зубов.
Первое звено в цепочке
В раннем эоцене, более 65 миллионов лет назад, жил первый прапредок современной лошади. Это «низкая лошадка» или Eohippus, которая была размером с собаку (до 30 см), опиралась на всю ступню конечности, на которой было по четыре (передние) и три (задние) пальца с маленькими копытцами. Питался эогиппус побегами и листьями и имел бугорчатые зубы. Буланая окраска и редкие волосы на подвижном хвосте - таков дальний предок лошадей и зебр на Земле.
Промежуточные звенья
Порядка 25 миллионов лет назад климат на планете изменился, и на смену лесам стали приходить степные просторы. В миоцене (20 миллионов лет назад) появляются мезогиппус и парагиппус, уже более похожие на современных лошадей. А первым травоядным предком в филогенетическом ряду лошади принято считать мерикгиппуса и плиогиппуса, которые выходят на арену жизни 2 миллиона лет назад. Гиппарион - последнее трехпалое звено
Этот предок жил в миоцене и плиоцене на равнинах Северной Америки, Азии и Африки. Эта трехпалая лошадка, напоминающая газель, еще не имела копыт, но могла быстро бегать, ела траву и именно она заняла огромные территории.

Однопалая лошадь - плиогиппус
Эти однопалые представители появляются 5 миллионов лет назад на тех же территориях, что и гиппарионы. Условия окружающей среды меняются - становятся еще суше, и степи значительно разрастаются. Вот тут и оказалась однопалость более важным признаком для выживания. Эти лошадки были высотой до 1,2 метра в холке, имели 19 пар ребер и сильные мышцы ног. Зубы у них приобретают длинные коронки и складки эмали с развитым цементным слоем.
Знакомая нам лошадь
Современная лошадь как конечный этап филогенетического ряда появилась в конце неогена, а в конце последнего ледникового периода (около 10 тысяч лет назад) в Европе и Азии уже паслись миллионы диких лошадей. Хотя усилия первобытных охотников и сокращение пастбищ сделали дикую лошадь редкостью уже 4 тысячи лет назад. Но два ее подвида - тарпан в России и лошадь Пржевальского в Монголии - сумели продержаться гораздо дольше всех остальных.

Дикие лошади
Сегодня настоящих диких лошадей практически не осталось. Российский тарпан считается вымершим видом, а лошадь Пржевальского в естественных условиях не встречается. Табуны лошадей, которые пасутся вольно, - это одичавшие одомашненные формы. Такие лошади хоть и быстро возвращаются к дикой жизни, но все же отличаются от истинно диких лошадей.
У них длинные гривы и хвосты, и они разномастные. Исключительно буланые лошади Пржевальского и мышастые тарпаны имеют как бы подстриженные челки, гривы и хвосты.

В Центральной и Северной Америке дикие лошади были полностью истреблены индейцами и появились там только после прибытия европейцев в XV веке. Одичавшие потомки лошадей конкистадоров дали начало многочисленным табунам мустангов, численность которых сегодня контролируется отстрелом.
Кроме мустангов в Северной Америке есть два вида диких островных пони - на островах Ассатиг и Сейбл. Полудикие табуны лошадей камаргу встречаются на юге Франции. В горах и болотах Британии тоже можно встретить некоторых диких пони.

Любимые наши скакуны
Человек приручил лошадь и вывел более 300 ее пород. От тяжеловесов до миниатюрных пони и красавцев скаковых пород. В России разводят около 50 пород лошадей. Самая известная из них - Орловская рысистая. Исключительно белая масть, отличная рысь и резвость - эти качества так ценил граф Орлов, которого считают основателем данной породы.
Классификация
Современная лошадь относится к семейству Лошадиные, куда кроме неё входят зебры и ослы. Семейство Лошадиные вместе с носорогами и тапирами принадлежат к отряду Непарнокопытные и являются потомками Condylarthra, от которой произошли все копытные. Группа Condylarthra - похожие на собак животные с пятипалыми конечностями, снабженными подушечками, давно вымершие примитивные млекопитающие древней палеоценовой группы.
Как протекала 60 миллионов лет назад эволюция лошади и как она постепенно приспосабливалась к изменяющимся условиям окружающей среды, ученые узнали благодаря изучению окаменелостей.
В это время они были совершенно не похожи на лошадь и больше напоминали динозавра. Наиболее древний предок лошади, останки которого сохранились до нас – барилямбда. Она была довольно толстой, длиной более двух метров, ноги её были короткими и заканчивались пятью пальцами с ногтями, отдалённо напоминающими копыто. Барилямбда питалась травой и мелкими кустарниками.
55 миллионов лет назад существовал континент, который можно назвать Евроамерикой. В то время современная Европа и Северная Америка еще не были разъединены Атлантическим океаном, а от Азии их отделял морской пролив, пролегавший там, где сейчас находится Уральский хребет. Это был цветущий континент, с тропическим климатом, покрытый вечнозелеными лесами. Воздух был полон густых дурманящих испарений, а земля взращивала богатый урожай кустарников, лиан и тропических великанов-деревьев, уходящих стволами далеко ввысь. Там, где через густой полог кроны тропического леса пробивались солнечные зайчики, на лесных полянах жили необычные, ростом с собаку, млекопитающие, питавшиеся листвой кустарников и мягкими лесными травами, - гиракотериумы, или эогиппусы (с лат. "лошади рассвета").
У них были маленькая голова на короткой шее, длинное туловище и короткие лапы. На передних ногах эогиппусы имели по четыре, а на задних - по три пальца. Мелкие зубы с низкими коронками были приспособлены для пережевывания только сочных мягких листьев да мелких беспозвоночных. Слабость жевательной системы подтверждалась и коротким лицевым отделом. Глазницы находились посредине черепа. Ноги у этого существа были полусогнутые и низкие. При ходьбе эогиппус опирался на подушечки пальцев, каждый из которых заканчивался коготком (будущие копыта). При передвижении животное напоминало современную собаку. Ясно, что с такими ногами нельзя было состязаться в скорости с быстроногими хищниками на открытых пространствах, поэтому эогиппус прятался в лесной чаще, зарослях кустарников и был «одет» в шкуру с коротким и редким волосяным покровом, полосатой или пятнистой окраски, подобно современным лесным и саванновым животным.

45 млн. лет назад жили маленькие, высотой менее полуметра лошадки – Орогиппусы.
Маленькие гривы на шеях этих лошадок были неподвижны, но зато хвосты, обросшие редкими волосами, развевались в воздухе. Их шерсть была буланой окраски, украшенная несколькими малозаметными продольными светлыми полосами. Передние ноги орогиппусов были ещё четырёхпалые, а задние - трёхпалые, и на всех пальцах были маленькие копытца. Но уже у этих первобытных лошадок кости средних пальцев ног были развиты сильнее, чем боковых.
Изменяется климат - меняются лошади
Останки процветавшего в эпоху олигоцена мезогиппуса свидетельствуют о существенных изменениях вида: ноги стали длиннее, спина выпрямилась (у эогиппуса спина была вогнутая), размеры животных стали крупнее. На передних ногах исчез один палец, и все четыре ноги стали трехпалыми. Изменились и зубы, малые коренные почти сравнялись с большими.

В миоцене произошло событие, значительно повлиявшее на развитие форм лошадиных. Климат на планете стал более континентальным, зима и лето обозначились гораздо четче, а тропики отступили к экватору: стали появляться безлесные пространства. Так как лесов стало меньше, то и часть животных вынуждена была поселиться на безлесных пространствах, а поселившись на них, вынуждена была как-то приспосабливаться к новым условиям. Примерно в это время возникают трехпалые миогиппусы, парагиппусы, мерикгиппусы и анхитерии.
"Когда трёхпалые анхитерии пошли на большие сухие луга древнего миоцена, то нужны были только ноги для опоры на сухой, твёрдой, невязкой почве; пошло развитие лошади и преобладание одного пальца..." - писал русский учёный Владимир Онуфриевич Ковалевский, которому мир обязан знаниями истории современной лошади. Однако это произошло не сразу. В новых условиях ноги у древних лошадей стали длиннее, и теперь животные могли пересекать большие пространства в поисках свежих и безопасных пастбищ.

На смену анхитерию пришел гиппарион - небольшая трехпалая лошадь, размером с осла, остатки которой обычно преобладают над другими ископаемыми. У него впервые из лошадей зубы обнаруживают приспособление для поедания трав. Поверхность зубов этого животного покрыта сложными эмалевыми петлями, препятствовавшими их истиранию при употреблении жесткой пищи. Вместе с тем зубы гиппариона еще имели более низкую коронку, чем у лошади, и, следовательно, были пригодны для истирания не очень жесткого корма. Строение конечностей гиппарионов говорит о том, что они были приспособлены для передвижения по мягким болотным почвам. Ноги гиппариона могли сгибаться под более острым углом, чем у лошади, так, как у северного оленя и лося, й, таким образом, при ходьбе и беге поднимались много выше, что позволяло ему легко передвигаться по высокотравью и кочковатым болотам.

Гиппарион представляет, по некоторым новым исследованиям, боковую ветвь, ныне вымершую, хотя в некоторых местностях существовавшую даже и одновременно с настоящими лошадьми. Гиппариона 2–3 миллиона лет назад заменила однопалая лошадь, возникшая от близкого рода плиогиппус, которая была лучше приспособлена к условиям жизни в степях и расселилась из Северной Америки по всем континентам.
Плиогиппусы были достаточно крупными, достигая в холке примерно 120 см, и жили, в отличие от своих предшественников, эогиппусов, в сухих степях. Копыта на средних пальцах плиогиппусов становились все более крупными и широкими. Они хорошо удерживали животных на твердой почве, давали им возможность разрывать снег, чтобы извлечь из-под него корм, защищаться от хищников, боковые же пальцы, наоборот, в ходе эволюции постоянно уменьшались, укорачивались так, что уже не касались земли, хотя были еще хорошо заметны, и в конце концов сохранились лишь в виде маленьких, похожих на стерженьки косточек прямо под кожей.

Такое изменение конечностей при эволюции лошади связано с тем, что потомки первобытных лошадок все чаще и чаще переселялись из болотистых и топких лесов на твердую почву сухих степей, заросших травой и кустарниками. В то время как для самых древних предков лошадей большее число пальцев на конечностях было оправданно, поскольку давало им большую безопасность при ходьбе по мягкой болотистой почве, жизнь в измененных условиях сделала более выгодным, чтобы боковые пальцы у них постепенно отмирали, а средние развивались, ведь в степи почва крепкая, твердая, пригодная не только для безопасной ходьбы, но и для стремительного галопа. Быстрое перемещение для плиогиппусов было жизненно важным, поскольку служило для них единственной защитой от нападения хищников.
Миллион лет назад появился настоящий Equus
В эпоху плейстоцена и ледниковых периодов Equus, прародитель современной лошади, по существовавшим в те времена перешейкам между материками мигрировал в Европу, Азию и Африку. Впоследствии, примерно десять тысяч лет назад, с окончанием оледенения эти перешейки (на их месте сейчас находятся такие проливы, как Гибралтарский и Берингов) исчезли, и это означало, что вымершие на одном из материков животные уже не могли вновь заселить его - по крайней мере без помощи человека. Именно это случилось в Америке: по неизвестной причине лошади там исчезли. Все члены современного семейства Equidae являются прекрасными бегунами и имеют на каждой ноге по одному функциональному пальцу (рудиментарные пальцы современной лошади - костяные наросты на тыльной стороне путового сустава - по-видимому, лишь напоминание о строении ступни эогиппуса). Все они ведут стадный образ жизни, и у всех коренные зубы приспособлены для перетирания травянистой пищи.

Каждая современная лошадь и пони по всем признакам ведет свой род от одного из трех типов, образовавшихся в различных климатических условиях. Северная Европа была родиной тяжелой, медленно передвигающейся лошади (Equus silvaticus), от которой ведут начало все известные породы тяжеловесов. Кроме нее в Азии существовала примитивная дикая лошадь, экземпляры которой были открыты в 1881 году (лошадь Пржевальского); а в Восточной Европе обитали более изящные тарпаны.
Потребовались долгие века, прежде чем живое существо в своём удивительном развитии прошло путь от маленького эогиппуса до современного коня - нашего самого благородного животного и верного помощника человека. Между ними - целая серия древних лошадей - от пятипалой лошадки размером с лисицу до европейского тарпана.
Палеонтологические данные по многим группам животных и некоторым группам растений выявили у них ряд морфологических признаков, изменяющихся на протяжении длительного периода времени в каком-то определённом направлении. Такие длительные изменения в одну сторону в пределах какой-либо естественной группы представляют собой эволюционные направления, существование которых факт, установленный палеонтологией. Об этом достаточно убедительно свидетельствуют также сравнительно-морфологические данные по многим современным группам растений, не представленным в палеонтологической летописи, но сохранившим ряд примитивных, промежуточных и продвинутых форм. Цель настоящей главы дать достаточно подробное описание одного хорошо документированного примера эволюционного направления, а также привлечь внимание к некоторым другим примерам. Наш подход к эволюционным направлениям носит здесь описательный, а не аналитический характер; природа эволюционных направлений будет рассмотрена в гл. 31 .
Примеры
Классическим примерам эволюционного направления служит изменение закрученности и скульптурированности раковины у пресноводной улитки Paludina (= Vivipara , Viviparidae ) в юго-восточной Европе в позднем плиоцене. Серии ископаемых улиток разного возраста были найдены в озерных отложениях в Югославии и на островах Эгейского моря. Серия раковин из Югославии показана на рис. 28.1. Она начинается предковой формой P. neumayri (рис. 28.1, neu) и доходит через промежуточные формы до P. hoernesi (рис. 28.1, hoer). С течением времени закрученность раковины становится более плотной, а её скульптурированность усложняется (Abel, 1929).
Плиоценовая P. neumayri близка к ныне живущему виду P. unicolor, который всё ещё встречается в Югославии и соседних областях. Производная форма P. hoernesi явно сильно отличается от предковой формы и её иногда выделяют в самостоятельный род Tulotoma . Её родичи больше не встречаются в Югославии, но представлены в современной фауне Северной Америки и Китая (Abel, 1929).
Другой классический пример увеличение рога у титанотерия (Brontotheriidae, Perissodactyla ) от эоцена до олигоцена (см. гл. 33 и рис. 33.2).
Некоторые направления эволюции наблюдаются одновременно в нескольких отдельных группах, развиваясь параллельно и независимо. Так, тенденция к увеличению размеров тела наблюдается у ряда наземных млекопитающих третичного периода, например у титанотериев, лошадей и хоботных; у динозавров мезозоя; у некоторых морских животных, например у морских улиток палеозоя. Тенденция к увеличению общих размеров, так широко распространенная среди животных, была названа правилом Копа (Соре).
Направления эволюции у цветковых растений обычно выявляют на основании сравнительно-морфологических, а не палеонтологических данных, поскольку первые в отличие от последних легко доступны (Cronquist, 1968; 1988; Тахтаджян, 1969; Stebbins, 1974; Carlquist, 1975).
У растений, так же как и у животных, нередко можно на блюдать параллельные эволюционные направления в несвязан ных между собой группах. Обычно это изменения в сторону уменьшения размеров и упрощения морфологии, известные под общим названием редукционных рядов. Редукционные ряды одного типа, наблюдаемые во многих независимых друг от друга группах, начинаются цветками с многочисленными лепестками, тычинками и (или) плодолистиками и кончаются цветками, имеющими лишь по нескольку этих органов. Другой обычный редукционный ряд от крупных многолетних до мелких однолетних трав. Часто такое направление у однолетних трав продолжается от перекрёстноопыляющихся форм с крупными цветками до самоопыляющихся форм с мелкими цветками.
Эволюционные ряды у Equidae
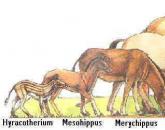
Самый знаменитый и наиболее хорошо документированный эволюционный ряд изменение признаков зубов, ног и других частей тела у Equidae в течение третичного периода. Очень полные палеонтологические данные по семейству лошадей охватывают период времени в 55 млн. лет, от раннего эоцена до настоящего времени. Краевые представители семейства это маленький четырёхпалый Hyracotherium (или Eohippus ) из верхнего палеоцена и эоцена и современный род Equus , к которому относятся лошади, дикие ослы, онагры и зебры. В палеонтологической летописи сохранился почти непрерывный ряд переходов, связывающих эти две формы.
Equidae происходят от ещё более древней палеоценовой группы Condylarthra , в которую входили небольшие, похожие на собак животные с пятипалыми конечностями, снабженными подушечками. Ряд ископаемых форм можно проследить ещё дальше вглубь от Hyracotherium до Condylarthra .
Филогения семейства лошадей была разработана на основании палеонтологических данных и хорошо известна вплоть до уровня родов. На рис. 28.2 изображена филогения этого семейства в сильно упрощенном виде. Геологический возраст и географическое распространение главных родов сем. Equidae также представлены на рис. 28.2. Эволюция этой группы происходила главным образом в Северной Америке, хотя в Евразии, Африке и Южной Америке в разное время в течение третичного и четвертичного периодов имела место эволюция боковых линий.
Главными направлениями эволюции лошадей были: 1) размеры тела; 2) длина хвоста; 3) механика конечности; 4) длина конечности; 5) форма головы; 6) величина головного мозга и степень его сложности; 7) коренные зубы. Нижеследующее изложение основано главным образом на работах Симпсона (Simpson, 1951; 1953) и Мак-Фаддена (MacFadden, 1985; 1988), где можно найти дополнительные детали.
Что касается размеров тела, то у Hyracotherium они были небольшими, как и у ранних Gondylarthra ; разные виды достигали 2550 см высоты в отличие от всем известных больших размеров Equus . Общее увеличение размеров тела происходило во многих линиях, в том числе у представителей Equinae из олигоцена и плиоцена. Об основных этапах этого эволюционного ряда позволяют судить изображения голов и конечностей, выполненные в одинаковом масштабе (рис. 28.3 и 28.5). Следует также отметить, что увеличение размеров, хотя оно достаточно хорошо выражено, происходило в разные периоды времени с различной скоростью, не распространялось на все семейство лошадей и нередко обращалось вспять у разных родов, в том числе и у самих Equus (Simpson, 1951; 1953; MacFadden, 1988).
Хвост изменялся от длинного по сравнению с размерами тела у Hyracotherium до относительно короткого и длинношерстного у Equus .
Наиболее ярко выраженным направлением в эволюции ноги лошади была редукция числа пальцев. Начальная точка этого направления предковые Condylarthra , которые были пятипалыми. У Hyracotherium на передних конечностях было по четыре, а на задних по три пальца (рис. 28.4, А), Лошади, жившие в олигоцене, миоцене и на протяжении большей части плиоцена, были трёхпалыми (рис. 28.4, Б и В). Среди трёхпалых лошадей можно заметить разницу между представителями более древних родов, у которых боковые пальцы хорошо развиты (рис. 28.4, Б), и более поздними формами, у которых они укорочены (рис. 28.4, В). Завершается этот ряд однопалой конечностью у Pliohippus , Hippidion и Equus (рис. 28.4, Г).
Уменьшение числа пальцев сопровождается изменением механики конечности. У Condylarthra и Hyracotherium на пальцах были подушечки, и эти животные при ходьбе опирались на эти подушечки, как это делают собаки. Самые древние трёхпалые лошади в олигоцене и некоторые из их прямых потомков в миоцене также опирались на подушечки (рис. 28.4, Б). Однако у другой ветви трёхпалых миоценовых лошадей и у более поздних однопалых лошадей нога обладает пружинящим действием. Масса тела ложится у них не на подушечки всех пальцев, которых уже нет, а на концевую фалангу среднего пальца, одетую копытом (рис. 28.3, В и Г; 28.4, В и Г).
По мнению Симпсона, у трёхпалых лошадей с пружинящей ногой два коротких боковых пальца, возможно, иногда служили для опоры, несколько облегчая нагрузку на средний палец (рис. 28.3, В и 28.4, В). У однопалых лошадей эти боковые пальцы в конце концов исчезли. У них масса тела целиком ложится на копыта, одевающие средние пальцы. Прочные связки, прикрепляющие мышцы к костям этих пальцев, играют роль рессор при беге.
Изменения в строении конечности носили эпизодический, а не медленный и постепенный характер; эти изменения происходили лишь в некоторых линиях и со сравнительно высокой скоростью.
Увеличение длины конечности в эволюции лошадей частично объясняется увеличением размеров тела. Переход от конечности, имеющей пальцы, к пружинящей ноге, снабженной копытом, ещё больше увеличил эффективную длину конечности, потому что при этом лошади стали ходить и бегать «на цыпочках».
На рис. 28.5 можно видеть многочисленные и сложные изменения как в пропорциях черепа, так и в его размерах. Морда вытягивается, а кости нижней челюсти становятся шире. Эти изменения в строении черепа связаны с изменениями зубов, которые будут рассмотрены в следующем разделе.
Другие изменения головы у Equidae коррелируют с изменением объема головного мозга. Hyracotherium отличался относительно небольшим мозгом по отношению к общим размерам (коэффициент энцефализации) (MacFadden, 1988). У олигоценового Mesohippus головной мозг также был довольно невелик. В дальнейшей истории семейства головной мозг увеличивался в размерах и становился сложнее по мере того, как в нем развивались области, связанные с развитием высшей нервной деятельности у современных млекопитающих. Прогрессивное развитие мозга и высшей нервной деятельности у лошадей происходило постепенно на протяжении всего третичного и четвертичного периодов.
Направления эволюции зубов у лошадей
Направления эволюции зубов у лошадей очень сложны и для их подробного описания пришлось бы прибегнуть к очень специальной терминологии. Для наших целей будет достаточно общего очерка эволюции зубов у лошадей с использованием минимального количества терминов.
Зубы Equus обладают следующими прогрессивными признаками: 1) дифференцировка режущих передних зубов для откусывания и коренных зубов для пережевывания; 2) большие размеры коренных зубов; 3) высокая коронка коренных зубов;
4) увеличение числа функциональных (жевательных) коренных зубов до шести на каждой стороне каждой челюсти; 5) сильное развитие на жевательной поверхности складок (лофодонтность), состоящих из эмали; 6) развитие между складками твердого цемента.
У Hyracotherium этим признакам соответствуют следующие примитивные особенности зубов: 1) более слабая дифференцировка передних и боковых зубов; 2) меньшие размеры зубов; 3) низкая коронка у коренных зубов; 4) отсутствие жевательных зубов (на каждой стороне каждой челюсти имелось по три коренных зуба, но это не были настоящие жевательные зубы);
5) складки эмали на жевательных поверхностях более простые и более низкие; 6) отсутствие цемента (рис. 28.5 и 28.6).
Эти различия в строении зубов отражают различия в пище. Hyracotherium был лесным животным, питавшимся листьями деревьев и кустарников, а быть может, и более разнообразной растительной пищей (MacFadden, 1988). Equus это, конечно, травоядное животное, обитавшее на лугах и в степях.
Многочисленные различия в признаках зубов между Equus и Hyracotherium возникли в результате ряда направлений эволюции; так, например, жевательные зубы стали крупнее (направление 2), коронки их стали выше (направление 3); складчатость усложнилась (направление 5) и т. д. Увеличение числа жевательных зубов произошло в результате постепенного превращения у Equus трёх ложнокоренных зубов Hyracotherium в зубы, сходные с коренными, процесс, известный под названием моляризации.
Эти изменения разных признаков зубов происходили не одновременно, а в разное время. Моляризация (направление 4) имела место главным образом в эоцене и олигоцене, а затем она более или менее прекратилась. Увеличение высоты коронки (направление 3) произошло в основном в миоцене, причём оно коснулось только таких вновь возникших травоядных групп, как Merychippus , тогда как у миоценовых лошадей, питавшихся листвой деревьев, коронки зубов оставались низкими.
Признаки зубов изменялись постепенно, один тип переходил в другой. Подобную интерградацию можно наблюдать в тех случаях, когда одна филетическая линия представлена ископаемыми зубами в ряде очень близких по возрасту геологических горизонтов. Рассмотрим линию ParahippusMerychippasNannippus из миоцена и плиоцена, изображенную на рис. 28.6. На рис. 28.7 показано изменение высоты коронки и жевательной поверхности в более полной последовательности стадий у той же линии. Ясно видно постепенное и почти непрерывное увеличение высоты коронки.
Далее можно дополнить позднемиоценовый отрезок ряда, изображенного на рис. 28.7. Даунс (Downs, 1961) провел подробное количественное изучение различий в зубах между двумя популяциями Merychippus из формаций Маскалл и Коалинга. Эти две популяции разделены промежутком в 12 млн. лет. Черты сходства и незначительные различия в складчатости зубов у этих популяций можно видеть на рис. 28.8.
Адаптивная природа направлений эволюции в семействе лошадей
Об адаптивном характере описанных выше изменений можно судить достаточно уверенно на основании тех условий, в которых жили животные. Важными факторами в эволюции лошадей, несомненно, были местообитание, пища и защита от врагов. Необходимо учитывать как прямые, так и косвенные воздействия этих и других факторов.
В начале третичного периода значительная часть суши была покрыта лесами, однако в результате изменений климата, происходивших в миоцене, эти обширные леса частично были замещёны открытыми пространствами саванн и прерий. В плиоцене площадь, занимаемая луговой растительностью, продолжала расширяться за счет лесов.
В начале третичного периода Equidae обитали в лесах и питались древесной листвой, как уже говорилось выше. Однако в миоцене одна из боковых линий семейства лошадей переселилась в открытые местообитания саванны и луга и перешла на питание травой. Такое изменение местообитания и характера питания произошло в Северной Америке; в результате эта боковая линия подверглась, вероятно, сильным новым давлениям отбора, воздействию которых можно приписать многие из наблюдаемых направлений эволюции.
Трава это грубая жесткая пища, от которой зубы стираются. Изменения зубов, происходившие у лошадей в миоцене и плиоцене, представляют собой по большей части первоначальные и усовершенствованные адаптации к питанию травой. Передние режущие зубы, длинные ряды коренных зубов и складки эмали и цемент на жевательных поверхностях этих зубов всё это необходимо для отщипывания и пережевывания травы. Высокие коронки коренных зубов увеличивают долговечность как самих этих зубов, так и их обладателей.
Некоторые изменения головы также, по-видимому, связаны с переходом к питанию травой. Удлинение морды способствует размещёнию двух дифференцированных наборов зубов резцов в передней части и коренных в задней. Широкие челюсти необходимы для того, чтобы в них могли сидеть коренные зубы с высокой коронкой; кроме того, челюсти должны быть мощными, чтобы пережевывать большие количества травы.
У Equus функции зубов тесно связаны с функциями специализированной пищеварительной системы. Эта пищеварительная система, характерными признаками которой являются наличие слепой кишки и быстрое прохождение пищи, приспособлена для переработки больших количеств грубоволокнистых трав и для извлечения достаточного количества питательных веществ из скуднойй низкокачественной растительности. Такие особенности Equus открывают для них степные местообитания. Equus способны выжить в голых степях, непригодных для большинства других копытных (Janis, 1976);
Животные, щиплющие траву на открытых пространствах саванн и равнин, гораздо более заметны для хищников, чем животные, обитающие в лесу. Увеличение размеров тела и большая сила один из эффективных способов защиты от хищников у наземных животных. Другой способ развитие высшей нервной деятельности, и, наконец, третий способность к быстрому бегу. Тенденции к развитию этих признаков в семействе лошадей, по всей вероятности, в значительной степени связаны с тем, что животные, обитающим на равнине, необходимы средства защиты.
Большие размеры тела позволяют решить некоторые проблемы, но создают и новые. Крупной лошади, питающейся травой, нужны для обеспечения питания более крупные, более твердые и более прочные зубы, чем маленькой лошади, также питающейся травой. Поэтому, как указывает Симпсон (Simpson, 1951), направления эволюции признаков зубов, вероятно, адаптивно коррелировали не только с изменением характера питания, но и с изменением размеров тела.
Увеличение размеров тела оказывает также побочное влияние на скорость бега. Крупной лошади труднее быстро бегать, чем маленькой, так как этому препятствует её тяжелое тело; для решения этой проблемы понадобилось усовершенствование механики конечности. Строение тела Hyracotherium было приспособлено для быстрого бега. Для того чтобы хотя бы сохранить такую скорость бега, которой обладал предковый Hyracotherium , не говоря уже о том, чтобы превысить её, более поздним крупным лошадям необходимо было изменить пропорции конечности и строение пальцев (Simpson, 1951).
Адаптивные аспекты правила Копа
Во многих группах животных наблюдается увеличение размеров тела в ходе филогенеза. Эта довольно обычная, но не всеобщая тенденция известна, как уже было отмечено, под названием правила Копа (Соре, 1896; Newell, 1949; Rensch, 1960; Stanley, 1973). Среди млекопитающих такое увеличение размеров наблюдается у сумчатых, хищных, непарнокопытных, парнокопытных, приматов и других групп; параллельные направления эволюции обнаружены у рептилий, членистоногих, моллюсков и в других крупных группах.
Большие размеры тела создают разнообразные селективные преимущества, во всяком случае у наземных животных. Здесь будут приведены четыре таких преимущества, к ним можно было бы добавить ряд других.
Размеры тела играют важную роль во взаимоотношениях хищник жертва. Хищник должен обладать определённой величиной, массой и силой, чтобы победить свою жертву; в свою очередь жертва должна иметь определённые размеры и силу, чтобы спастись от хищника. Многие сочетания хищникжертва оказываются втянутыми в своего рода «гонку вооружений», ведущую ко всё большему возрастанию размеров тела у обоих партнеров. Возможно, что между лошадьми и крупными хищниками сложились именно такие взаимоотношения и с такими результатами.
Для развития высшей нервной деятельности необходим большой и сложный головной мозг, а для такого мозга нужна большая голова, что в свою очередь требует большого тела. Отбор на повышение уровня развития мозга мог повлечь за собой отбор на увеличение размеров тела как коррелятивного признака.
Сила, необходимая для борьбы, ещё один признак, коррелирующий с размерами тела. В драках за обладание самками, которые происходят между самцами у многих видов копытных, превосходство в размерах и силе даёт селективное преимущество. Некоторые направления эволюции, касающиеся изменения размеров, могут быть отчасти результатом полового отбора. У крупных животных есть ещё одно преимущество над мелкими, связанное с сохранением тепла в организме. Большие тела теряют меньше тепла, чем небольшие, вследствие более благоприятного соотношения между площадью поверхности и объемом.
Тенденция к увеличению размеров никогда не была всеобщей. В некоторых филетических линиях наблюдается такое увеличение, тогда как в других родственных им линиях на протяжении того же периода отмечается, напротив, уменьшение размеров. Известны и такие случаи, когда в одной и той же филетической линии в один период времени эволюция была направлена в сторону увеличения размеров, а в другой в сторону их уменьшения. Оба эти отклонения от правила Копа обнаружены в семействе лошадей, а также у слонов, оленей и в других группах.
Все это свидетельствует об относительности селективного преимущества, создаваемого большими размерами тела. Увеличение этих размеров выгодно, но лишь в известных условиях. В других условиях оно может не давать никаких преимуществ или даже оказаться невыгодным. У островных животных в общем отмечается тенденция к уменьшению размеров по сравнению с их материковыми предками, что представляет собой отчасти результат отбора на способность к выживанию при строго ограниченных пищевых ресурсах (примеры см. Carlquist, 1965).
Макроэволюция - эволюционные преобразования, происходящие на надвидовом уровне: роды, семейства, отряды, классы, типы. Термин "макроэволюция" в биологию ввел в 1927 г. ученый-селекционер Ю. А. Филипченко.
Макроэволюция - это процесс формирования крупных систематических групп (тип, класс, отряд).
Палеонтология - наука, изучающая ископаемые остатки организмов. Ученые-налеонтологи, опираясь на идеи Дарвина, исследовали историю развития животных. В. О. Ковалевский (1842- 1883) в основном занимался исследованием парнокопытных животных. Он, изучив причины эволюционного процесса на примере лошади, сделал вывод о том, что изменения в ее строении связаны с изменением среды обитания (рис. 27). С изменением климата уменьшались лесные массивы, а увеличение открытых степных пространств способствовало изменению жизненной среды предков лошадей. Они стали приспосабливаться к условиям степей. Для спасения от врагов и поиска пастбищ с богатой растительностью необходимо было научиться быстро бегать. Перемещения на большие расстояния способствовали изменению числа пальцев на конечностях. Изменениям подверглись не только конечности, но и форма тела, черепа, строение зубов - в связи с пережевыванием травы. Произошла полная перестройка организма лошади. Такими изменениями явились: уменьшение количества пальцев от пяти до одного, укрепление тела, усложнение коренных зубов. Все это свидетельствует о процессе эволюции лошади. Итак, постепенные переходы одних видов в другие, одних родов в другие - доказательство эволюционного процесса.
Рис. 27. Эволюция лошади
В. О. Ковалевский изучал эволюцию современных лошадей и назвал ступени их развития названиями веков третичного периода кайнозойской эры. Лошадей расположил по мере уменьшения числа пальцев на конечности.
1. Фенакодус (существовал в палеоцене), пятипалый, ростом с лисицу.
2. Эогиппус (жил в эоцене), рост 30 см, с четырехпалыми передними конечностями, трехпалыми задними.
3. Миогиппус (жил в миоцене), хорошо развит средний палец, второй и четвертый пальцы короткие.
4. Парагиппус (жил в миоцене), сильно развит средний палец, второй и четвертый пальцы укорочены.
5. Плиогиппус (существовал в плиоцене), однопалый, остальные пальцы атрофированы.
6. Современная лошадь.
Открытие В. О. Ковалевским систематического ряда в изучении видов ископаемых лошадей - типичный пример эволюционного процесса. Наличие последовательно сменяющих друг друга форм позволяет построить ряд с высокой степенью достоверности эволюции, который называется филогенетическим.
В результате изучения ископаемых остатков парнокопытных млекопитающих В. О. Ковалевский и его преемники выяснили историю развития лошади (филогенез).
Макроэволюция. Палеонтология. Палеонтологические доказательства. Филогенетический ряд.
Макроэволюция - эволюционные преобразования, происходящие на надвидовом уровне.
Достижения палеонтологии в изучении истории развития лошади.
Филогенетический ряд доказывает существование эволюции.
1. Что такое макроэволюция и лшкроэволюция? В чем их отличие?
2. Что изучает палеонтология?
1. Что способствовало изменению жизненной среды предков лошадей?
2. Кто исследовал эволюцию лошадей и на какие материалы он опирался?
1. Опишите схематично изменения в эволюции лошади.
2. Дайте описание предков лошадей, живших в каждом веке.
Происхождение первых лошадей
Непарнокопытные (отряд класса млекопитающих животных) одарили человечество только двумя видами домашних животных: лошадью и ослом. Оба из семейства лошадиных. Осёл стал домашним раньше лошади, примерно 6 тысяч лет назад в Египте. Предком его был африканский дикий осёл, ныне уцелевший лишь кое-где в Эфиопии и Сомали. Он находится там под защитой закона, запрещающего убивать диких ослов.
Лошадь одомашнена лишь в третьем тысячелетии до нашей эры. Прародиной её были южнорусские степи, а диким предком - тарпан. В древности в Нижней Месопотамии и в Вавилонии приручались и азиатские дикие ослы. Их запрягали в телеги, использовали как вьючных животных. Но с появлением лошади домашние азиатские ослы стали довольно быстро исчезать. А, кроме того, их вытесняли и более «мощные», более пригодные для тяжёлых работ домашние ослы Африки, вскоре появившиеся в Азии.
Если бы человек увидел в лесу эогиппуса, то никогда бы не подумал, что это предок нашей лошади. Ростом он был не больше лисицы. Голову имел маленькую, шею короткую, спину горбатую, шкуру полосатую, а лапы четырёхпалые (передние) и трёхпалые (задние). Жил эогиппус в сырых лесах Северной Америки 50 миллионов лет назад. Питался листьями. Было несколько разновидностей эогиппусов, некоторые из них рано переселились в Европу (по-видимому, через «мост», существовавший тогда на севере между Канадой, Гренландией, Исландией и Скандинавией). Европейский потомок эогиппуса - палеотерий могучим телосложением напоминал носорога.
Первым лошадям в Европе не повезло, здесь они все вымерли. Но в Америке род их по-прежнему процветал. От эогиппуса произошёл орогиппус, а от него трёхпалый мезогиппус, который был уже ростом с овцу. Тут в истории Земли произошло важное событие: сырые тропические леса, покрывавшие большую часть планеты, стали всюду исчезать. Появились степи и луговые травы. Мезогиппусы вышли из лесных за рослей и рискнули начать новую жизнь под открытым небом прерий. Питаться стали травой.

В степи их преследовали быстроногие предки волков. Спасение было только в одном: научиться бегать быстрее хищников. Лишние пальцы на ногах стали обузой (на одном пальце бегать легче!), и по ископаемым костям видно, как у предков лошадей стал атрофироваться палец за пальцем, пока на каждой ноге не осталось лишь по одному. Лошадь превратилась в однокопытное животное.
Но это случилось не сразу. От мезогиппуса произошёл меригиппус, а затем стройный гиппарион (ростом чуть пониже зебры). Два недоразвитых боковых пальца на его ногах не касались земли. Трёхпалый гиппарион бегал, следовательно, уже на одном пальце.
Едва ли какое-нибудь другое копытное животное встречалось такими колоссальными стадами, как гиппарион. Миллионные полчища этих элегантных лошадей через перешеек, соединявший в те времена Чукотку и Аляску, проникли из Северной Америки в Азию, а затем и в Европу. Бесчисленные табуны гиппарионов галопировали по равнинам Евразии.

Их ископаемые остатки так многочисленны, что палеонтологи назвали «фауной гиппариона» весь комплекс живых существ, обитавших в тех же степях в одно время с этими лошадьми. В Африку, Южную Америку и Австралию гиппарионы не сумели пробраться: тогда эти страны отделялись от Северной Америки, Азии и Европы широкими проливами и морями. Прошло несколько миллионов лет, и все гиппарионы вымерли.
Более счастливая судьба ожидала двоюродного, так сказать, «брата» гиппариона (конечно, в эволюционном, а не бытовом смысле) - плиогиппуса. От него-то и произошли наши лошади. Когда-то табуны плиогиппусов населяли всю Северную и Южную Америку, Европу, Азию и Африку (к тому времени эти материки снова соединились перешейками).
Среди древних лошадей были очень интересные разновидности: одни больше самого крупного тяжеловоза, другие меньше карликового пони. Но миллион лет назад все лошади в Америке по непонятной причине вымерли. В Африке уцелели лишь зебры и ослы, а в Европе и Азии - два диких вида, история которых отныне тесно сплетена с судьбой человека.
В ледниковое время, несколько десятков тысяч лет назад, дикие лошади водились ещё по всей Европе. Вместе с мамонтами и северными оленями они часто попадали на обед к троглодитам, первобытным людям, жившим в пещерах. О том свидетельствуют «кухонные» отбросы наших предков - огромные кучи раздробленных костей, исследованные антропологами. В одной из них нашли остатки десяти тысяч съеденных лошадей. Прародители наши, как видно, не страдали отсутствием аппетита.



Популярное
- Сценарий музыкальной сказки «Лягушка-путешественница» по мотивам сказки Песня «Испекли бы пироги»
- Лошадь знак зодиака. Лошадь. Восточный китайский гороскоп для Лошади Год лошади какие бывают мужчины
- Миопатия у ребенка 1 год
- Физическая подготовка является
- Упражнения для похудения рук и плеч в домашних условиях
- Подвижные игры, включающие бег, прыжки, метание Игры – эстафеты
- Мок назвал критерии допуска российских спортсменов на олимпиаду
- Висы и упоры в гимнастике 4 кл
- Книга: Беспощадная истина - Майк Тайсон
- Стиль боксирования и техника мэнни пакьяо Последний бой пакьяо